지방산 대사
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
지방산 대사는 지방산의 분해와 합성을 아우르는 생화학적 과정이다. 지방산은 트리글리세리드 형태로 저장되며, 에너지가 필요할 때 리파아제에 의해 분해되어 혈액으로 운반되어 대사 세포로 유입된다. 세포 내에서 아실-CoA로 활성화된 지방산은 카르니틴 셔틀을 거쳐 미토콘드리아에서 베타 산화를 통해 아세틸-CoA로 분해되어 시트르산 회로와 전자전달계를 통해 ATP를 생성한다. 지방산 생합성은 팔미트산이 생성될 때까지 반복적인 과정을 거치며, 지방산 대사는 호르몬과 알로스테릭 조절을 받는다. 지방산 대사 이상은 고지혈증과 같은 질환을 유발할 수 있으며, 암세포의 지방산 대사 변화도 연구되고 있다. 지방산은 세포막 구성, 신호 전달, 에이코사노이드 합성 등 다양한 기능을 수행하며, 프로스타글란딘과 같은 생리활성 물질의 전구체로 작용한다.

더 읽어볼만한 페이지
- 간장학 - 간세포암
간세포암은 간에서 발생하는 악성 종양으로, 만성 간 질환 환자에게서 주로 발생하며, 다양한 원인과 위험 요인에 의해 발생하고, 복통, 황달 등의 증상이 나타날 수 있으며, 수술, 간 이식 등의 치료 방법이 사용되고, 예방을 위해 B형 간염 예방 접종 등이 중요하다. - 간장학 - 담관
담관은 간에서 십이지장으로 연결되는 관 네트워크로, 담즙을 운반하여 지방의 소화와 흡수를 돕고, 폐쇄 시 황달을 유발하며, 담관 배액 시술 등으로 치료한다. - 지방산 - 발레르산
발레르산은 5개의 탄소 원자를 가진 카복실산으로 불쾌한 냄새가 나는 무색 액체이며, 휘발성 에스터 제조에 사용되어 향수, 화장품, 식품 등에 활용되고 발레리안 뿌리 등에서 발견된다. - 지방산 - 아라키드산
아라키드산은 탄소 20개로 이루어진 포화 지방산으로, 땅콩 기름과 같은 식물성 기름에 많이 함유되어 있으며, 인체 내에서 세포막 구성, 신호 전달 물질의 전구체, 에이코사노이드 합성에 관여하고, 제약 산업에서 프로스타글란딘의 전구체로 사용되는 경제적 가치가 높은 물질이다. - 물질대사 - 아라키드산
아라키드산은 탄소 20개로 이루어진 포화 지방산으로, 땅콩 기름과 같은 식물성 기름에 많이 함유되어 있으며, 인체 내에서 세포막 구성, 신호 전달 물질의 전구체, 에이코사노이드 합성에 관여하고, 제약 산업에서 프로스타글란딘의 전구체로 사용되는 경제적 가치가 높은 물질이다. - 물질대사 - 카프릴산
카프릴산은 8개의 탄소로 이루어진 포화 지방산으로, 식품, 향료, 소독제 등 다양한 산업에 사용되며, 식욕 촉진, 항균 작용, 간질 치료 등 의학적 용도로도 연구된다.
2. 지방산 이화작용
지방산 대사는 크게 지방산 이화작용과 지방산 생합성으로 나뉜다. 지방산 이화작용은 지방산을 분해하여 에너지를 생성하는 과정이며, 지방산 생합성은 반대로 에너지를 사용하여 지방산을 합성하는 과정이다.
지방산 이화작용은 β 산화 과정을 통해 아세틸-CoA를 생성하고, 이는 시트르산 회로로 들어가 에너지를 생성한다.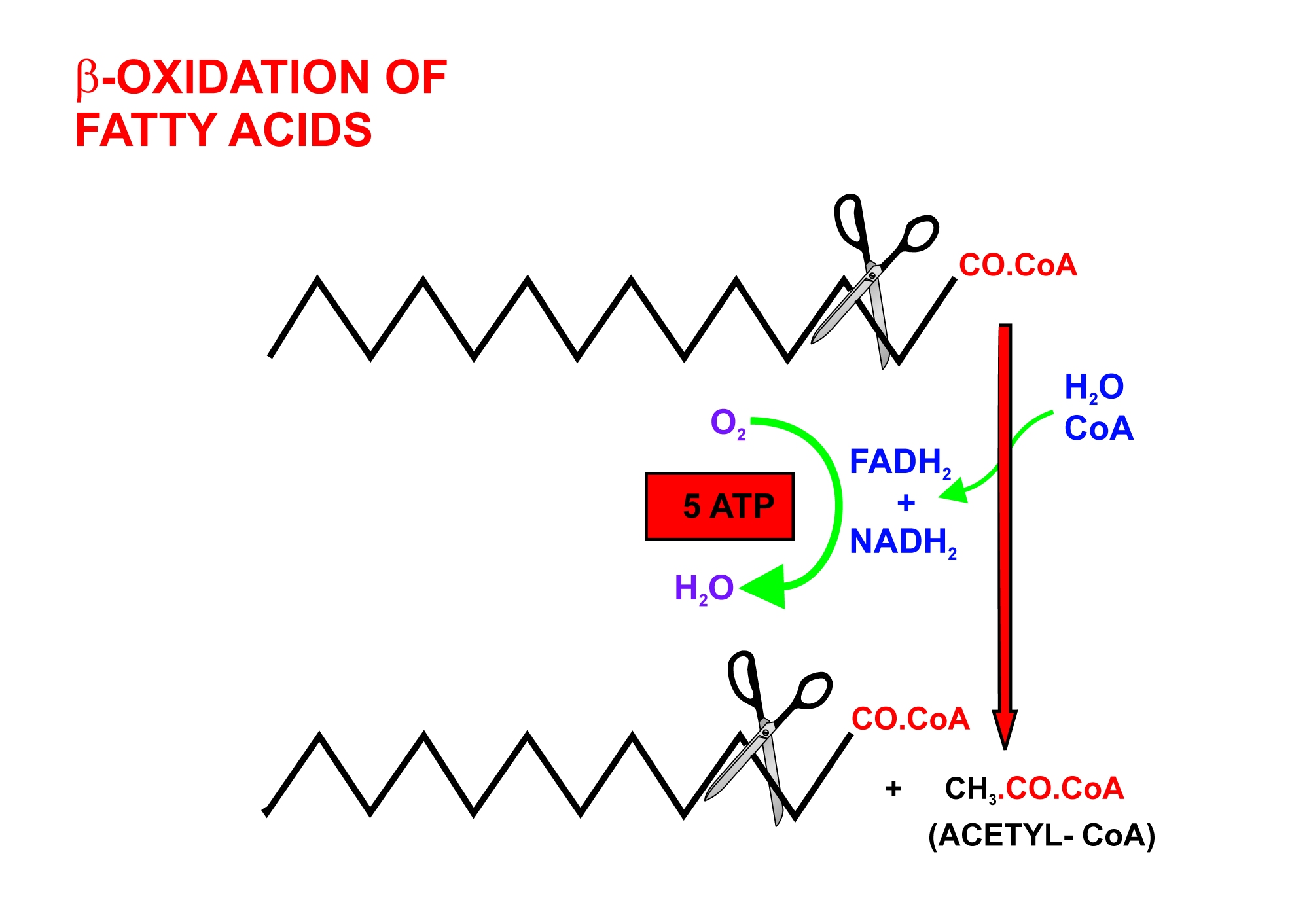
지방산은 트리글리세리드 형태로 지방 조직에 저장되었다가, 필요에 따라 리파아제에 의해 지방 분해되어 유리지방산 형태로 혈액을 통해 이동한다. 유리지방산은 세포 내로 유입된 후 아실-CoA로 활성화되어 미토콘드리아로 이동하여 β 산화 과정을 거친다.
베타 산화의 각 단계는 다음과 같다.[2]
1. 아실-CoA 탈수소효소에 의한 탈수소화 (1 FADH2 생성)
2. 에노일-CoA 수화효소에 의한 수화
3. 3-히드록시아실-CoA 탈수소효소에 의한 탈수소화 (1 NADH + H+ 생성)
4. 티올라아제에 의한 절단 (1 아세틸-CoA 및 2개의 탄소로 짧아진 지방산 생성)
이 과정은 지방산이 완전히 아세틸-CoA로 분해될 때까지 반복된다. 홀수 탄소 지방산의 경우 최종적으로 아세틸-CoA와 프로피오닐-CoA가 생성된다. 각 베타 산화 과정에서 5 ATP가 생성된다.[14][15]
아세틸 CoA는 시트르산 회로에서 옥살아세트산과 결합하여 시트르산을 형성하고, 산화적 인산화를 통해 이산화탄소(CO2)와 물(H2O)로 완전히 산화되면서 1 GTP 및 11 ATP를 생성한다.[2][10] 프로피오닐-CoA는 여러 단계를 거쳐 숙시닐-CoA로 전환된 후 시트르산 회로로 들어간다.[16][17]
간에서는 특정 상황(단식, 기아, 저탄수화물 식단, 장기간의 격렬한 운동, 조절되지 않는 1형 당뇨병)에서 옥살아세트산이 포도당신생합성에 사용될 수 있다. 이 경우 아세틸-CoA는 아세토아세트산, 베타-히드록시부티르산 등의 케톤체를 형성한다.[10] 케톤체는 혈액을 통해 다른 조직으로 운반되어 에너지원으로 사용될 수 있으며, 특히 혈액-뇌 장벽을 통과하여 뇌의 에너지원으로 사용될 수 있다.[10]
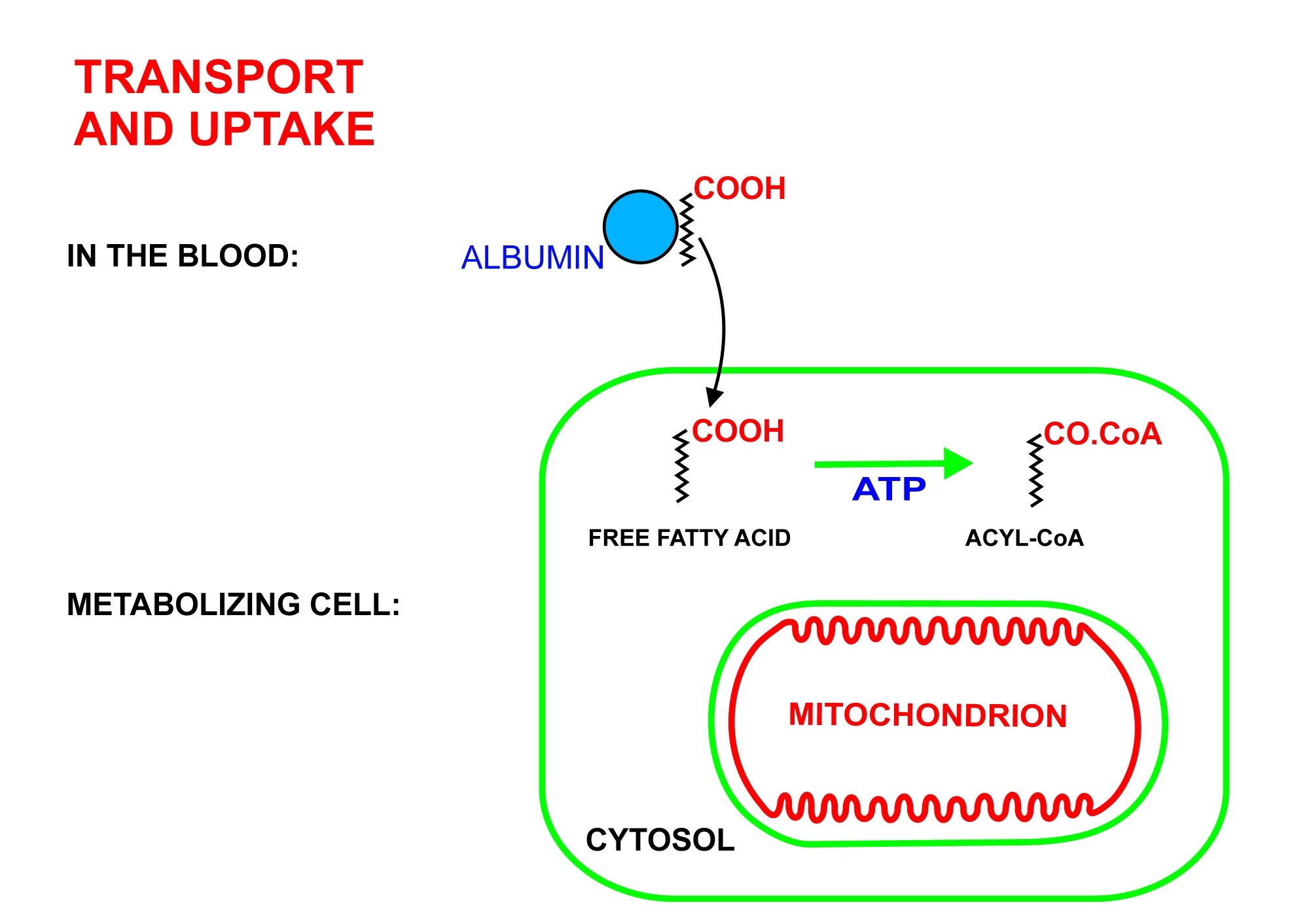
2. 1. 지방산의 세포 내 유입 및 활성화
지방산은 트리글리세리드 형태로 지방 조직의 지방 저장소에 저장된다. 식사 사이에는 다음과 같이 지방산이 방출된다.- 리파아제는 지방 분해를 수행하여 지방산 사슬을 트리글리세리드 형태로 저장된 글리세롤에서 제거한다. 이러한 리파아제는 식사 후 혈액 글루코스 수치가 감소하여 혈액 내 높은 에피네프린 및 글루카곤 수치(또는 지방 조직의 교감 신경에서 분비되는 노르에피네프린)에 의해 활성화되며, 동시에 혈액 내 인슐린 수치를 낮춘다.[1]
- 글리세롤에서 해리되면 유리지방산은 혈액으로 들어가 혈장 알부민에 부착되어 신체 전체로 수송된다.[4]
- 장쇄 유리지방산은 특정 수송 단백질을 통해 대사 세포(즉, 적혈구 및 중추 신경계의 뉴런을 제외한 신체의 대부분의 생존 세포)로 들어간다. 예를 들어 SLC27 계열 지방산 수송 단백질이 있다.[5][6] 적혈구는 미토콘드리아를 포함하지 않으므로 지방산을 대사할 수 없으며, 중추 신경계 조직은 미토콘드리아를 포함하고 있음에도 불구하고 장쇄 지방산(중쇄 지방산과 반대로[7][8])이 혈액-뇌 장벽[9]을 통과하여 이러한 세포를 적시는 간질액으로 들어갈 수 없기 때문에 지방산을 사용할 수 없다.
- 세포 내에서 장쇄 지방산—CoA 연결효소는 지방산 분자와 ATP (AMP와 무기 피로인산으로 분해됨) 사이의 반응을 촉매하여 지방 아실-아데닐레이트를 생성한 다음 자유 조효소 A와 반응하여 지방 아실-CoA 분자를 생성한다.
- 아실-CoA가 미토콘드리아로 들어가기 위해서는 카르니틴 셔틀이 사용된다.[10][11][12]
1. 아실-CoA는 미토콘드리아 외막 및 미토콘드리아 내막의 세포질 측면에 위치한 카르니틴 팔미토일전이효소 I에 의해 카르니틴의 히드록실기로 전이된다.
2. 아실-카르니틴은 카르니틴이 외부로 이동함에 따라 카르니틴-아실카르니틴 전위효소에 의해 내부로 이동한다.
3. 아실-카르니틴은 미토콘드리아 내막의 내부 측면에 위치한 카르니틴 팔미토일전이효소 II에 의해 아실-CoA로 다시 전환된다. 방출된 카르니틴은 세포질로 다시 이동하고, 아실-CoA는 미토콘드리아 기질로 이동한다.
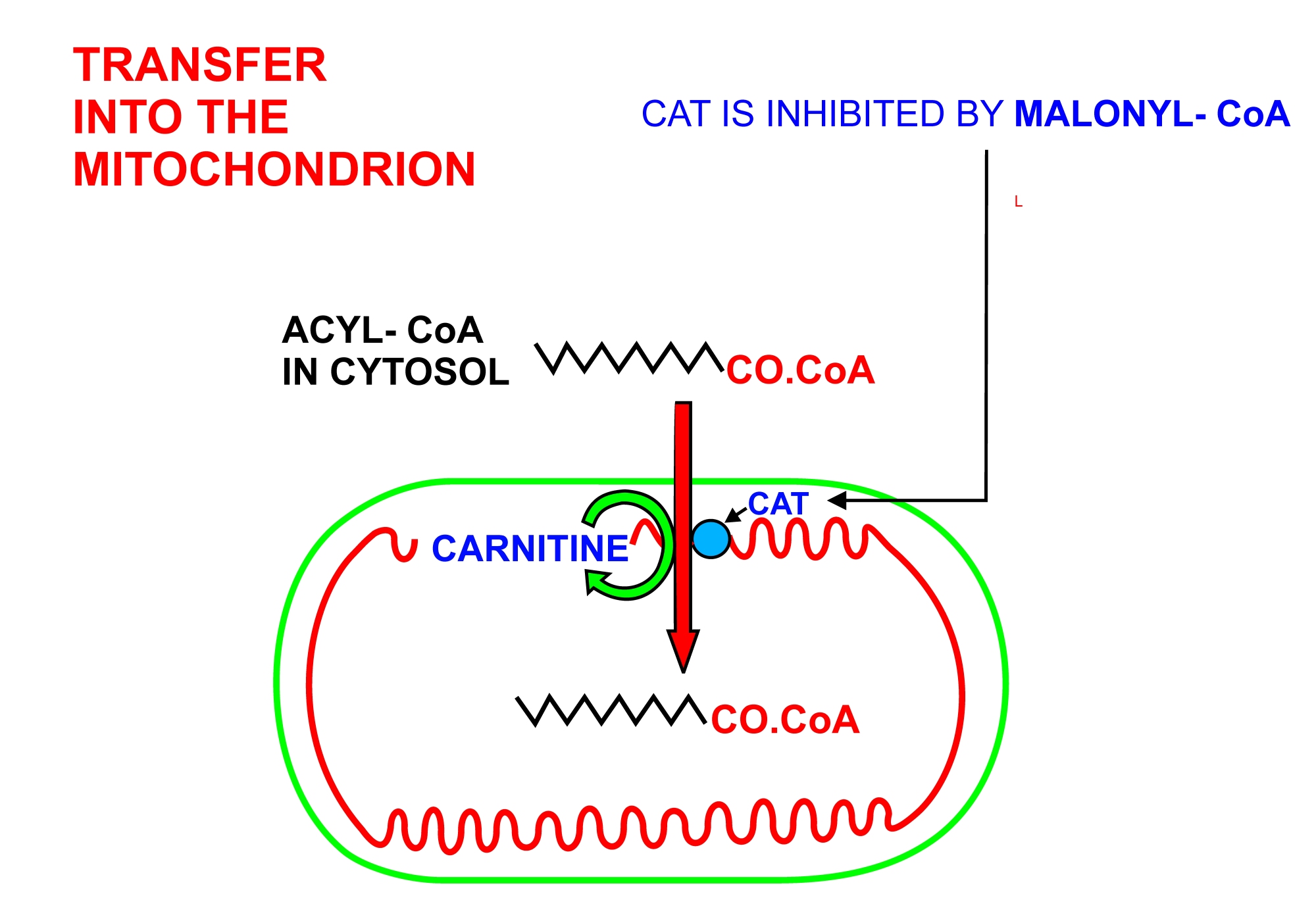
2. 2. 아실-CoA의 미토콘드리아 내 이동
아실-CoA가 미토콘드리아로 들어가기 위해서는 카르니틴 셔틀이 사용된다.[10][11][12]# 아실-CoA는 미토콘드리아 외막 및 미토콘드리아 내막의 세포질 측면에 위치한 카르니틴 팔미토일전이효소 I에 의해 카르니틴의 하이드록실기로 전이된다.
# 아실-카르니틴은 카르니틴이 외부로 이동함에 따라 카르니틴-아실카르니틴 전위효소에 의해 내부로 이동한다.
# 아실-카르니틴은 미토콘드리아 내막의 내부 측면에 위치한 카르니틴 팔미토일전이효소 II에 의해 아실-CoA로 다시 전환된다. 방출된 카르니틴은 세포질로 다시 이동하고, 아실-CoA는 미토콘드리아 기질로 이동한다.
2. 3. 베타 산화 (β-oxidation)
지방산은 트리글리세리드 형태로 지방 조직의 지방 저장소에 저장된다. 식사 사이에는 다음과 같이 방출된다.- 리파아제는 지방 분해를 수행하여 지방산 사슬을 트리글리세리드(또는 지방) 형태로 저장된 글리세롤에서 제거한다. 이러한 리파아제는 식사 후 혈액 글루코스 수치가 감소하여 혈액 내 높은 에피네프린 및 글루카곤 수치(또는 지방 조직의 교감 신경에서 분비되는 노르에피네프린)에 의해 활성화되며, 동시에 혈액 내 인슐린 수치를 낮춘다.[1]
- 글리세롤에서 해리되면 유리지방산은 혈액으로 들어가 혈장 알부민에 부착되어 신체 전체로 수송된다.[4]
- 장쇄 유리지방산은 특정 수송 단백질을 통해 대사 세포(즉, 적혈구 및 중추 신경계의 뉴런을 제외한 신체의 대부분의 생존 세포)로 들어간다. 예를 들어 SLC27 계열 지방산 수송 단백질.[5][6] 적혈구는 미토콘드리아를 포함하지 않으므로 지방산을 대사할 수 없으며, 중추 신경계 조직은 미토콘드리아를 포함하고 있음에도 불구하고 장쇄 지방산(중쇄 지방산과 반대로[7][8])이 혈액-뇌 장벽[9]을 통과하여 이러한 세포를 적시는 간질액으로 들어갈 수 없기 때문에 지방산을 사용할 수 없다.
- 세포 내에서 장쇄 지방산—CoA 연결효소는 지방산 분자와 ATP (AMP와 무기 피로인산으로 분해됨) 사이의 반응을 촉매하여 지방 아실-아데닐레이트를 생성한 다음 자유 조효소 A와 반응하여 지방 아실-CoA 분자를 생성한다.
- 아실-CoA가 미토콘드리아로 들어가기 위해서는 카르니틴 셔틀이 사용된다.[10][11][12]
1. 아실-CoA는 미토콘드리아 외막 및 미토콘드리아 내막의 세포질 측면에 위치한 카르니틴 팔미토일전이효소 I에 의해 카르니틴의 히드록실기로 전이된다.
2. 아실-카르니틴은 카르니틴이 외부로 이동함에 따라 카르니틴-아실카르니틴 전위효소에 의해 내부로 이동한다.
3. 아실-카르니틴은 미토콘드리아 내막의 내부 측면에 위치한 카르니틴 팔미토일전이효소 II에 의해 아실-CoA로 다시 전환된다. 방출된 카르니틴은 세포질로 다시 이동하고, 아실-CoA는 미토콘드리아 기질로 이동한다.
- 베타 산화는 미토콘드리아 기질에서 아실-CoA 분자 형태의 지방산의 긴 탄소 사슬을 일련의 2탄소(아세테이트) 단위로 절단하며, 이는 조효소 A와 결합하여 아세틸 CoA 분자를 형성하며, 이는 시트르산 회로의 "시작" 부분에서 옥살아세테이트와 축합하여 시트르산을 형성한다.[13] 이 반응은 연료 - 아세틸-CoA -가 사이클에 추가되어 각 사이클 회전 및 후속 산화적 인산화 과정에서 CO2 및 H2O로 소실되고 ATP 형태로 포착된 상당량의 에너지가 방출되므로 사이클의 "시작점"을 표시하는 것으로 생각하는 것이 편리하다.
베타 산화의 단계는 다음과 같다.[2]
1. 아실-CoA 탈수소효소에 의한 탈수소화, 1 FADH2 생성
2. 에노일-CoA 수화효소에 의한 수화
3. 3-히드록시아실-CoA 탈수소효소에 의한 탈수소화, 1 NADH + H+ 생성
4. 티올라아제에 의한 절단, 1 아세틸-CoA 및 2개의 탄소로 짧아진 지방산(새로운, 짧아진 아실-CoA 형성) 생성
이 베타 산화 반응은 지방산이 완전히 아세틸-CoA로 환원될 때까지 또는 탄소 원자 수가 홀수인 지방산의 경우 지방산 분자당 아세틸-CoA 및 1 프로피오닐-CoA 분자까지 반복된다. 아실-CoA 분자의 각 베타 산화 절단은 결국 산화적 인산화 과정에서 5 ATP 분자를 생성한다.[14][15]
- 베타 산화에 의해 생성된 아세틸-CoA는 시트르산 회로의 옥살아세테이트와 결합하여 시트르산을 형성함으로써 미토콘드리아 내에서 시트르산 회로로 들어간다. 산화적 인산화와 결합하면 아세틸-CoA가 CO2와 물로 완전 연소된다. 이 과정에서 방출된 에너지는 아세틸-CoA 분자당 1 GTP 및 11 ATP 분자 형태로 포착된다.[2][10] 이것은 간에서 특정 상황을 제외하고 지방산의 베타 산화가 발생하는 곳마다 아세틸-CoA의 운명이다.
프로피오닐-CoA는 나중에 생물질-의존성 프로피오닐-CoA 카르복실라아제 (PCC) 및 비타민 B12-의존성 메틸말로닐-CoA 뮤테이즈 (MCM)를 통해 순차적으로 숙시닐-CoA로 전환된다.[16][17] 숙시닐-CoA는 먼저 말산으로 전환된 다음 피루브산으로 전환된 후 기질로 이동하여 시트르산 회로로 들어간다.
2. 4. 시트르산 회로 및 전자전달계를 통한 ATP 생성
지방산은 트리글리세라이드 형태로 유기체에 저장되며, 산소가 적고 무수물이므로 농축된 음식 에너지의 에너지원이다. 지방산 1 그램당 에너지 수율은 탄수화물의 4kcal(17kJ)보다 훨씬 높은 약 9 kcal(37kJ)이다. 지방산의 탄화수소 부분은 소수성이므로 비교적 무수 환경(무수 환경)에 저장될 수 있다. 반면에 탄수화물은 수화도가 더 높다. 예를 들어 1g의 글리코겐은 약 2g의 물과 결합하며, 이는 1.33kcal/g(4kcal/3g)으로 변환된다. 즉, 지방산은 저장된 질량 단위당 6배 이상의 에너지를 보유할 수 있다.[10]동면하는 동물은 지방 저장고를 연료로 사용하는 좋은 예이다. 예를 들어, 곰은 약 7개월 동안 동면하며, 이 전체 기간 동안 에너지는 지방 저장고의 분해에서 얻어진다. 철새는 마찬가지로 대륙 간 여행을 시작하기 전에 많은 지방 저장고를 축적한다.[18]
젊은 성인 인간의 지방 저장량은 평균 약 10~20kg이지만, 성별과 개인적인 성향에 따라 크게 다르다.[19] 반대로, 인체는 약 400g의 글리코겐만 저장하며, 이 중 300g은 골격근 내에 잠겨 있어 신체 전체에서 사용할 수 없다. 간에 저장된 약 100g의 글리코겐은 단식 1일 이내에 고갈된다.[10] 그 후, 간에서 신체 조직의 일반적인 사용을 위해 혈액으로 방출되는 포도당은 글루코겐 아미노산과 몇 가지 다른 포도당 신생 기질로부터 합성되어야 하며, 여기에는 지방산이 포함되지 않는다.[1] 그럼에도 불구하고, 지방분해는 글리세롤을 방출하며, 이는 포도당 신생 경로로 들어갈 수 있다.
2. 5. 케톤체 생성
간에서는 포도당신생합성 경로를 통해 옥살로아세테이트가 전부 또는 부분적으로 전환될 수 있는데, 이는 단식, 기아, 저탄수화물 식단, 장기간의 격렬한 운동 및 조절되지 않는 1형 당뇨병 상황에서 일어난다. 이러한 상황에서 옥살로아세테이트는 말산으로 수소화된 후 간세포의 미토콘드리아에서 세포질로 이동하여 포도당으로 전환되고 혈액으로 방출된다.[10] 혈중 낮은(또는 없는) 인슐린과 높은 글루카곤 농도에 의해 포도당신생합성이 크게 자극되면, 간에서 옥살로아세테이트는 아세틸-CoA와 축합할 수 없게 된다. 대신 아세틸-CoA는 아세토아세트산 및 베타-히드록시부티르산을 형성하도록 전환된다.[10] 아세토아세트산, 베타-히드록시부티르산, 그리고 이들의 자발적 분해 생성물인 아세톤은 종종 케톤체로 불리는데, 이는 엄밀히 말하면 "체"가 아닌 수용성 화학 물질이다. 생성된 케톤체는 간에서 혈액으로 방출된다.미토콘드리아를 가진 모든 세포는 혈액에서 케톤체를 흡수하여 시트르산 회로에서 연료로 사용할 수 있는 아세틸-CoA로 다시 전환할 수 있다. 다른 조직들은 간에서처럼 옥살로아세테이트를 포도당신생합성 경로로 전환할 수 없기 때문이다. 유리지방산과 달리 케톤체는 혈액-뇌 장벽을 통과할 수 있어, 중추 신경계 세포가 일반적으로 사용하는 포도당을 대체하여 연료로 사용될 수 있다.[10] 단식, 저탄수화물 식단, 장기간의 격렬한 운동 또는 조절되지 않는 1형 당뇨병에서 혈액 내 케톤체 농도가 높아지는 현상을 케토시스라 하며, 극단적인 형태로는 조절되지 않는 1형 당뇨병에서 케토산증으로 나타난다.
3. 지방산 생합성
지방산 생합성은 에너지를 축적하는 반응으로, FAS(Fatty acid synthase)가 관여한다. 아세틸-CoA가 아세틸 CoA 카복실레이스(ACC, acetyl CoA carboxylase) 효소에 의해 말로닐-CoA로 전환되면서 시작된다.[1] 지방산 생합성은 미토콘드리아 외부의 세포질에서 아세틸-CoA로부터 합성된다. 지방산 생합성 경로는 지방산 분해 경로 (베타 산화)와 억제적인 관계에 있다.
베타 산화와 마찬가지로, 직선형 지방산 생합성은 16개의 탄소를 가진 팔미트산이 생성될 때까지 6가지 반복 반응을 통해 일어난다.[35][36] 위의 그림은 미생물에서 지방산이 어떻게 합성되는지 보여주며, 대장균에서 발견되는 효소를 나열한다.[35] 이러한 반응은 일반적으로 하나의 복합체로 작용하는 여러 효소를 포함하는 지방산 합성효소 II (FASII)에 의해 수행된다. FASII는 원핵생물, 식물, 곰팡이, 기생충뿐만 아니라 미토콘드리아에도 존재한다.[37]
동물과 효모와 같은 일부 곰팡이에서 이러한 반응은 지방산 합성효소 I (FASI)에서 발생한다. FASI는 지방산을 생성하는 데 필요한 모든 효소 활성을 가진 큰 이량체 단백질이다. FASI는 FASII보다 덜 효율적이지만, 초기 사슬 종결을 통해 "중간 사슬" 지방산을 포함한 더 많은 분자를 형성할 수 있게 한다.[37]
지방산 합성 중 환원제는 NADPH인 반면, NAD는 베타 산화 (아세틸-CoA로의 지방산 분해)에서 산화제이다. NADPH는 생합성 반응 동안 소비되는 반면 NADH는 에너지 생성 반응에서 생성된다는 일반적인 원리를 보여준다.[41] NADPH는 오탄당 인산 경로 또는 사과산이 피루브산으로 전환될 때 생성된다.[41]
인간의 경우, 지방산은 주로 간과 지방 조직, 그리고 수유 중인 유선에서 탄수화물로부터 생성된다. 해당과정에 의해 생성된 피루브산은 탄수화물을 지방산과 콜레스테롤로 전환하는 중요한 중간체이다.[41]
3. 1. 아세틸-CoA의 세포질 이동
지방산은 미토콘드리아 내부에서 베타 산화를 통해 아세틸-CoA로 분해되지만, 지방산 합성은 미토콘드리아 외부의 세포질에서 아세틸-CoA로부터 일어난다. 이 두 경로는 장소, 반응, 기질에서 뚜렷하게 구분되며 서로 억제적인 관계를 가진다.[1] 피루브산 탈수소효소 복합체 반응은 비가역적이기 때문에, 베타 산화로 생성된 아세틸-CoA는 피루브산으로 전환될 수 없다.[10] 대신 옥살로아세트산과 결합하여 시트르산 회로로 들어간다.인간의 경우, 지방산은 주로 간과 지방 조직, 수유 중인 유선에서 탄수화물로부터 생성된다. 해당과정으로 생성된 피루브산은 미토콘드리아 내에서 아세틸-CoA로 전환되어 탄수화물을 지방산과 콜레스테롤로 전환하는 중요한 중간체 역할을 한다.[41] 그러나 지방산과 콜레스테롤 합성은 세포질에서 일어나기 때문에 아세틸-CoA는 세포질로 수송되어야 한다. 이는 직접적으로 일어날 수 없다.
세포질 아세틸-CoA를 얻기 위해, 시트르산 회로에서 구연산 (아세틸-CoA와 옥살로아세트산의 축합으로 생성)이 제거되어 미토콘드리아 내막을 통해 세포질로 운반된다.[41] 세포질에서 ATP 구연산 분해 효소에 의해 아세틸-CoA와 옥살로아세트산으로 분해된다. 옥살로아세트산은 말산으로 전환되어 미토콘드리아로 돌아가고, 다시 옥살로아세트산으로 전환되어 더 많은 아세틸-CoA를 미토콘드리아 밖으로 수송한다.[42] 세포질 아세틸-CoA는 아세틸-CoA 카복실화 효소에 의해 말로닐-CoA로 카복실화되며, 이는 지방산 합성의 첫 번째 단계이다.[42][43]
3. 2. 말로닐-CoA 생성
지방산 생합성은 아세틸-CoA가 아세틸 CoA 카복실레이스(ACC) 효소에 의해 말로닐-CoA로 전환되면서 시작된다.[35]3. 3. 지방산 합성효소 복합체 (Fatty Acid Synthase, FAS)
FAS(Fatty acid synthase)는 지방산 생합성에 관여하는 효소 복합체이다. 지방산 생합성은 에너지를 축적하는 반응으로, 아세틸-CoA가 아세틸 CoA 카복실레이스(ACC, acetyl CoA carboxylase)에 의해 말로닐-CoA로 전환되면서 시작된다.[35][36]베타 산화와 마찬가지로, 직선형 지방산 생합성은 16개의 탄소를 가진 팔미트산이 생성될 때까지 6가지 반복 반응을 통해 일어난다.[35]
위의 그림은 미생물에서 지방산이 어떻게 합성되는지를 보여주며, 대장균에서 발견되는 효소를 나열하고 있다.[35] 이러한 반응은 일반적으로 하나의 복합체로 작용하는 여러 효소를 포함하는 지방산 합성효소 II (FASII)에 의해 수행된다. FASII는 원핵생물, 식물, 곰팡이, 기생충 및 미토콘드리아에 존재한다.[37]
동물과 효모와 같은 일부 곰팡이에서는 이러한 반응이 지방산 합성효소 I (FASI)에서 발생한다. FASI는 지방산을 생성하는 데 필요한 모든 효소 활성을 가진 큰 이량체 단백질이다. FASI는 FASII보다 덜 효율적이지만, 초기 사슬 종결을 통해 "중간 사슬" 지방산을 포함한 더 많은 분자를 형성할 수 있게 한다.[37]
효소, 아실전이효소 및 트랜스아실라제는 아실 수용체와 공여체 사이에서 지방산을 전달하여 인지질, 트리아실글리세롤 등에 지방산을 포함시킨다. 또한 생리활성 지질과 그 전구체 분자를 합성하는 역할도 한다.[38]
16:0 탄소 지방산이 형성되면, 탈포화 및/또는 연장을 초래하는 여러 가지 변형을 겪을 수 있다. 스테아르산(18:0)으로 시작하는 연장은 여러 막 결합 효소에 의해 주로 소포체에서 수행된다. 연장 과정과 관련된 효소 단계는 기본적으로 지방산 생합성에 의해 수행되는 단계와 동일하지만, 연장의 네 가지 주요 연속 단계는 물리적으로 연관될 수 있는 개별 단백질에 의해 수행된다.[39][40]






약어: ACP – 아실 운반 단백질, CoA – 조효소 A, NADP – 니코틴아미드 아데닌 디뉴클레오티드 인산
지방산 합성 중 환원제는 NADPH인 반면, 베타 산화(아세틸-CoA로의 지방산 분해)에서 산화제는 NAD이다. NADPH는 생합성 반응 동안 소비되는 반면 NADH는 에너지 생성 반응에서 생성된다.[41] NADPH의 공급원은 두 가지이다. 말산이 "NADP+-연결 말산 효소"에 의해 산화적 탈카르복실화되면 피루브산, CO2 및 NADPH가 생성된다. NADPH는 또한 포도당을 리보스로 변환하는 오탄당 인산 경로에 의해 형성되며, 이는 뉴클레오타이드와 핵산의 합성에 사용되거나 피루브산으로 대사될 수 있다.[41]
3. 4. 지방산의 변형
베타 산화와 마찬가지로, 직선형 지방산 생합성은 16개의 탄소 팔미트산이 생성될 때까지 6가지 반복 반응을 통해 일어난다.[35][36]제시된 그림은 미생물에서 지방산이 어떻게 합성되는지 보여주며, 대장균에서 발견되는 효소를 나열한다.[35] 이러한 반응은 일반적으로 하나의 복합체로 작용하는 여러 효소를 포함하는 지방산 합성효소 II (FASII)에 의해 수행된다. FASII는 원핵생물, 식물, 곰팡이, 기생충뿐만 아니라 미토콘드리아에도 존재한다.[37]
동물과 효모와 같은 일부 곰팡이에서 이러한 동일한 반응은 지방산을 생성하는 데 필요한 모든 효소 활성을 가진 큰 이량체 단백질인 지방산 합성효소 I (FASI)에서 발생한다. FASI는 FASII보다 덜 효율적이지만, 초기 사슬 종결을 통해 "중간 사슬" 지방산을 포함한 더 많은 분자를 형성할 수 있게 한다.[37]
16:0 탄소 지방산이 형성되면, 탈포화 및/또는 연장을 초래하는 여러 가지 변형을 겪을 수 있다. 스테아르산(18:0)으로 시작하는 연장은 여러 막 결합 효소에 의해 주로 소포체에서 수행된다. 연장 과정과 관련된 효소 단계는 기본적으로 지방산 생합성에 의해 수행되는 단계와 동일하지만, 연장의 네 가지 주요 연속 단계는 물리적으로 연관될 수 있는 개별 단백질에 의해 수행된다.[39][40]
아래의 표는 지방산 변형 단계를 보여준다.
약어: ACP – 아실 운반 단백질, CoA – 조효소 A, NADP – 니코틴아미드 아데닌 디뉴클레오티드 인산.
지방산 합성 중 환원제는 NADPH인 반면, NAD는 베타 산화 (아세틸-CoA로의 지방산 분해)에서 산화제이다. 이러한 차이는 NADPH가 생합성 반응 동안 소비되는 반면 NADH는 에너지 생성 반응에서 생성된다는 일반적인 원리를 보여준다.[41]
효소, 아실전이효소 및 트랜스아실라제는 아실 수용체와 공여체 사이에서 지방산을 전달하여 인지질, 트라이아실글리세롤 등에 지방산을 포함시킨다. 또한 생리활성 지질과 그 전구체 분자를 합성하는 역할도 한다.[38]
4. 지방산 대사의 조절
지방산 대사는 β 산화 과정처럼 에너지를 방출하는 반응과 FAS가 관여하는 에너지 축적 반응으로 나뉜다. β 산화 과정을 거쳐 생성된 아세틸-CoA는 시트르산 회로로 들어가고, 지방산 생합성 과정은 아세틸-CoA가 아세틸 CoA 카복실레이스(ACC)에 의해 말로닐-CoA로 전환되며 시작된다.
MAPK/ERK 경로는 포도당과 젖산 농도를 조절 인자로 사용하여, 골격근 근섬유와 근원섬유 세포 내에서 느리지만 지속적인 근육 활동을 감지하면, 빠른 에너지를 제공하는 포도당보다 저장된 지방산을 분해하여 에너지원으로 사용하도록 신호전달체계를 작동시킨다.
아세틸-CoA는 아세틸-CoA 카복실화 효소에 의해 말로닐-CoA로 형성되어 지방산 합성 경로에 투입된다. 아세틸-CoA 카복실화 효소는 포화 직쇄 지방산 합성의 조절 지점이며, 인산화와 알로스테릭 조절을 받는다. 인산화 조절은 주로 포유류에서, 알로스테릭 조절은 대부분의 생물체에서 일어난다. 알로스테릭 조절은 팔미토일-CoA에 의한 피드백 억제와 시트르산에 의한 활성화로 발생한다. 포화 지방산 합성의 최종 생성물인 팔미토일-CoA 농도가 높으면, 세포 내 지방산 축적을 방지하기 위해 아세틸-CoA 카복실화 효소를 알로스테릭하게 비활성화시킨다. 시트르산은 높은 수준에서 아세틸-CoA 카복실화 효소를 활성화시키는데, 이는 크레브스 회로에 투입되어 에너지를 생산할 수 있는 충분한 아세틸-CoA가 있음을 의미한다.[44]
혈장 내 인슐린 농도가 높으면(예: 식사 후) 아세틸-CoA 카복실화 효소의 탈인산화를 유발하여 활성화되고, 아세틸-CoA로부터 말로닐-CoA 형성을 촉진하여 탄수화물을 지방산으로 전환한다. 반면, 에피네프린과 글루카곤(기아 및 운동 중 혈액으로 방출됨)은 이 효소의 인산화를 유발하여 지방 생성을 억제하고 베타 산화를 통한 지방산 산화를 선호한다.[41][43]
5. 지방산 대사 이상
지방산 대사 장애는 고트리글리세리드혈증(트리글리세리드 수치가 너무 높은 경우) 또는 다른 유형의 고지혈증으로 설명할 수 있다. 이러한 장애는 유전적이거나 후천적일 수 있다.
지방산 대사 장애의 유전적 유형은 일반적으로 지질 대사 이상으로 분류된다. 이러한 장애는 지방산 산화 장애 또는 ''지질 저장 질환''으로 설명될 수 있으며, 근육, 간 및 기타 세포 내에서 에너지를 생산하기 위해 신체가 지방산을 산화하는 능력에 영향을 미치는 효소 또는 수송 단백질 결함으로 인해 발생하는 여러 가지 선천성 대사 이상 중 하나이다. 지방산 산화 장애가 근육에 영향을 미치는 경우, 이는 대사성 근병증이다.
또한 암세포는 지방산 합성[45] 및 미토콘드리아 지방산 산화(FAO)[46] 모두에 관해 불규칙한 지방산 대사를 보일 수 있으며, 이는 종양 발생 및 세포 성장의 다양한 측면에 관여한다.
6. 기타 지방산의 기능
MAPK/ERK 경로는 포도당과 젖산 농도를 조절 인자처럼 사용하여, 골격근의 근섬유와 근원섬유 세포 내에서 느리지만 지속적인 근육 활동을 감지한다. 이에 따라 빠른 에너지를 제공하는 포도당보다 저장된 지방산을 분해하여 에너지원으로 사용하도록 하는 신호 전달 체계를 작동시킬 수 있다고 알려져 있다.[25]
6. 1. 신호 전달
MAPK/ERK 경로는 포도당과 젖산의 농도를 조절 인자처럼 사용하여, 골격근의 근섬유와 근원섬유 세포 내에서 느리지만 지속적인 근육 활동을 감지한다. 이에 따라 빠른 에너지를 제공하는 포도당보다 저장된 지방산을 분해하여 에너지원으로 사용하도록 하는 신호 전달 체계를 작동시킬 수 있다고 알려져 있다.[25]
지방산은 세포의 세포막을 구성하는 인지질의 필수적인 부분이다. 이러한 인지질은 세포막 결합 효소 인산화효소 C(PLC)에 의한 인지질 포스파티딜이노시톨 4,5-이중인산(PIP)의 가수분해를 통해 디아실글리세롤(DAG)과 이노시톨 삼인산(IP)으로 분해될 수 있다.[25]
6. 2. 에이코사노이드 합성


지방산 대사의 산물 중 하나는 동물에서 다양한 호르몬과 유사한 효과를 나타내는 화합물인 프로스타글란딘이다. 프로스타글란딘은 사람과 다른 동물의 거의 모든 조직에서 발견된다. 이들은 20개의 탄소로 구성된 다불포화 지방산인 아라키돈산으로부터 효소적으로 유도된다. 따라서 모든 프로스타글란딘은 20개의 탄소 원자를 포함하며, 5개의 탄소 고리를 포함한다. 이들은 에이코사노이드의 하위 분류이며 지방산 유도체의 프로스타노이드 클래스를 형성한다.[26]
프로스타글란딘은 세포막을 구성하는 인지질로부터 아라키도네이트를 절단하여 합성된다. 이는 막 인지질에 직접 작용하는 포스포리파제 A2에 의해, 또는 DAG(디아실글리세롤)에 작용하는 리파제에 의해 촉매된다. 그런 다음 아라키도네이트는 프로스타글란딘 합성효소의 사이클로옥시게나제 성분에 의해 작용한다. 이는 지방산 사슬의 중간 부분에 대략 사이클로펜탄 고리를 형성한다. 또한 이 반응은 두 분자의 O2에서 유래된 4개의 산소 원자를 첨가한다. 생성된 분자는 프로스타글란딘 G2이며, 이 분자는 효소 복합체의 하이드로퍼옥시다제 성분에 의해 프로스타글란딘 H2로 전환된다. 이 매우 불안정한 화합물은 다른 프로스타글란딘, 프로스타사이클린 및 트롬복산으로 빠르게 변환된다.[26] 그런 다음 이들은 에이코사노이드 호르몬을 제조한 세포를 둘러싼 간질액으로 방출된다.
아라키도네이트가 사이클로옥시게나제 대신 리폭시게나제에 의해 작용하면 하이드록시에이코사테트라에노산과 류코트리엔이 형성된다. 이들 또한 국소 호르몬으로 작용한다.
프로스타글란딘은 두 가지 유도체인 프로스타사이클린과 트롬복산을 갖는다. 프로스타사이클린은 국소적으로 작용하는 강력한 혈관확장제이며 혈액 혈소판의 응집을 억제한다. 혈관 확장의 역할 덕분에 프로스타사이클린은 염증에도 관여한다. 이들은 혈관 벽에서 합성되며 불필요한 혈전 형성을 방지하고 평활근 조직의 수축을 조절하는 생리적 기능을 한다.[27] 반대로, 트롬복산(혈소판 세포에서 생성)은 혈관수축제이며 혈소판 응집을 촉진한다. 이들의 이름은 혈전 형성(혈전증)에서의 역할에서 유래되었다.
참조
[1]
서적
In: Biochemistry.
W.H. Freeman and Company
1995
[2]
웹사이트
Oxidation of fatty acids
http://pharmaxchange[...]
[3]
논문
Lipolysis: pathway under construction
[4]
웹사이트
Mobilization and cellular uptake of stored fats (triacylglycerols) (with animation)
http://pharmaxchange[...]
[5]
논문
A current review of fatty acid transport proteins (SLC27)
2004-02-01
[6]
논문
SLC27 fatty acid transport proteins
2013-04
[7]
논문
Energy contribution of octanoate to intact rat brain metabolism measured by 13C nuclear magnetic resonance spectroscopy
2003-07
[8]
논문
Heptanoate as a neural fuel: energetic and neurotransmitter precursors in normal and glucose transporter I-deficient (G1D) brain
2013-02
[9]
서적
In: Biochemistry.
W.H. Freeman and Company
1995
[10]
서적
Biochemistry.
W.H. Freeman and Company
1995
[11]
웹사이트
Activation and transportation of fatty acids to the mitochondria via the carnitine shuttle (with animation)
http://pharmaxchange[...]
[12]
논문
l-Carnitine Supplementation in Childhood Epilepsy: Current Perspectives
[13]
웹사이트
Oxidation of fatty acids
http://pharmaxchange[...]
[14]
웹사이트
Oxidation of odd carbon chain length fatty acids
http://pharmaxchange[...]
[15]
웹사이트
Oxidation of unsaturated fatty acids
http://pharmaxchange[...]
[16]
논문
Propionyl-CoA carboxylase - A review
2017-12
[17]
논문
Comparative aspects of propionate metabolism
[18]
서적
Biochemistry.
W.H. Freeman and Company
1995
[19]
논문
Body composition work capacity and work efficiency of active and inactive young men
1973
[20]
논문
Toxicology, metabolism, and biochemistry of 1,2-propanediol
[21]
논문
You Can Get There From Here: Acetone, Anionic Ketones and Even-Carbon Fatty Acids can Provide Substrates for Gluconeogenesis
https://www.ajol.inf[...]
2016-08-07
[22]
논문
Metabolite and isotopomer balancing in the analysis of metabolic cycles: II. Applications
http://web.mit.edu/b[...]
[23]
논문
Propanediol metabolism and its relation to lactic acid metabolism
[24]
논문
Reduction of trioses by NADPH-dependent aldo-keto reductases. Aldose reductase, methylglyoxal, and diabetic complications
[25]
서적
In: Biochemistry.
W.H. Freeman and Company
1995
[26]
서적
In: Biochemistry.
W.H. Freeman and Company
1995
[27]
서적
An introduction to behavioral endocrinology
Sinauer Associates
[28]
웹사이트
Digestion of fats (triacylglycerols)
http://pharmaxchange[...]
[29]
논문
The function of bile salts in fat absorption. The solvent properties of dilute micellar solutions of conjugated bile salts
1963
[30]
서적
In: Biochemistry.
W.H. Freeman and Company
1995
[31]
서적
Advanced nutrition and human metabolism
Wadsworth/Cengage Learning
2013
[32]
서적
In: Gray's Anatomy
Churchill Livingstone
1989
[33]
서적
In: Biochemistry.
W.H. Freeman and Company
1995
[34]
논문
Insulin-induced dephosphorylation of hormone-sensitive lipase
[35]
간행물
Fatty Acid Biosynthesis
Blackwell Pub.
2008
[36]
웹사이트
MetaCyc pathway: superpathway of fatty acids biosynthesis
http://www.biocyc.or[...]
BioCyc
[37]
서적
AOCS Lipid Library
https://lipidlibrary[...]
2011-04-20
[38]
간행물
Acyltransferases and transacylases that determine the fatty acid composition of glycerolipids and the metabolism of bioactive lipid mediators in mammalian cells and model organisms
2014-01-01
[39]
웹사이트
MetaCyc pathway: stearate biosynthesis I (animals)
http://www.biocyc.or[...]
BioCyc
[40]
웹사이트
MetaCyc pathway: very long chain fatty acid biosynthesis II
http://www.biocyc.or[...]
BioCyc
[41]
서적
Biochemistry.
W.H. Freeman and Company
1995
[42]
간행물
SREBP-1c Transcription Factor and Lipid Homeostasis: Clinical Perspective
http://content.karge[...]
2007
[43]
서적
Fundamentals of Biochemistry, 2nd Edition
https://archive.org/[...]
John Wiley and Sons, Inc.
[44]
웹사이트
Fatty Acid Synthesis
http://rpi.edu/dept/[...]
[45]
간행물
Clinical importance of FASN in relation to HIF-1α and SREBP-1c in gastric adenocarcinoma
https://www.scienced[...]
2019-05
[46]
간행물
Downregulation of fatty acid oxidation by involvement of HIF-1α and PPARγ in human gastric adenocarcinoma and its related clinical significance
https://pubmed.ncbi.[...]
2021-05
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com